Mathematical Modeling in
Computational Biology:
Focus on Excitable Media and
Cardiac Electrophysiology
Modeling normal human ventricular cell and
tissue electrophysiology. I have been involved in developing a minimal model of human
ventricular cell electrophysiology based closely on published data for three
anatomical regions of the ventricular wall (epicardial, endocardial, and
midmyocardial) that experience gradients in certain electrophysiological
properties. Previously, three other models human ventricular cell physiology
had been published, none of which adequately matched newly obtained
experimental data. In addition, the other existing models do not provide
accurate descriptions of the regional gradients across the ventricular wall.
Using the minimal model, my colleagues and I have been able to explore the
contribution of these regional gradients to the morphological characteristics
of the electrocardiogram (ECG) using a realistic three-dimensional ventricular
anatomical model and specifically to match experimental data obtained using a
ventricular slab similar to experimental setups (see Fig. 1).

Figure 1. (A) Action potential duration
(APD) as a function of diastolic interval (DI) obtained from experiments (dots)
and using the minimal model (solid) for epicardial (blue), endocardial (green),
and midmyocardial (red) human ventricular cells, with corresponding simulated
action potentials. (B) Anatomical ventricular slab matching experimental slab
conditions including regional variations in electrophysiological properties in
the epicardium (blue), midmyocardium (red), and endocardium (green), with
corresponding action potentials. (C) Simulated ECG in the anatomical structure
shown in (B) obtained using the minimal model.
Modeling Brugada Syndrome. I have been developing a model of
diseased physiology reflecting differences believed to be present with a type
of genetic disease called the Brugada syndrome, which can cause sudden cardiac
death without any underlying structural defects. In many cases, Brugada
Syndrome has been shown to occur in the presence sodium ion channel mutation
that results in loss of function that decreases sodium current. Most
experimental data on Brugada syndrome has been obtained using dogs, so I
initially developed a model to reproduce important properties of these
experiments. More recently, Brugada syndrome data in humans has become
available, and accordingly I have been working to implement these
characteristics within the context of the human ventricular cell model. Among
other things, I have been able to demonstrate the importance of regional
variations in cell properties in inducing reentry reentrant arrhythmias (see
Fig. 2) and have identified the conditions necessary for a reentrant arrhythmia
(spiral wave) to occur, including slow pacing rates and appropriate sodium
channel recovery from inactivation. Following this preliminary work, it is
still necessary to investigate how Brugada Syndrome arrhythmias develop in the
full anatomical context of the human ventricles, complete with anatomical
complexity and regional gradients in ion channel properties.

Figure 2. Arrhythmia development in
an anatomically realistic three-dimensional model of the canine right
ventricular free wall due to the presence of Brugada syndrome, which causes
various regions of the epicardium to delay the action potential dome (A,
lower left) or to lose the dome entirely (A, upper left) compared to the normal
epicardial action potential (A, right). Because
repolarization time varies by region (B-C), the late dome region is able to
generate a new wave that propagates into the fully repolarized loss-of-dome
region above it (D-E). This initiates a reentrant arrhythmia (F-G).
Electrical wave fronts are shown in red, with quiescent tissue in dark blue.
{kind=link}
Movies of Brugada arrhythmia development in 2D preparations with the same distribution of normal, late dome, and lost dome regions as in Fig. 2A. Stimulation from the left and from below result in arrhythmia initiation, while stimulation from the right does not induce an arrhythmia.
{kind=link}
{kind=link}
{kind=link}
Modeling rabbit ventricular cell and tissue
electrophysiology. To
understand arrhythmia dynamics in rabbit ventricles, mathematical
it was necessary to construct models of rabbit ventricular action potentials to match
electrophysiological properties in the presence of two different motion
uncoupling agents used for optical mapping experiments: diacetyl
monoxime and cytochalasin-D.
These pharmacological agents are used to prevent ventricular contractions that
would interfere with image recording. However, they also affect
electrophysiological properties. Starting from action potential recordings and restitution
information recorded in tissue using both optical and microelectrode
techniques, two models were developed and then simulated in two- and
three-dimensional structures. In two-dimensional sheets, spiral waves initiated
using either model remained stable with linear core trajectories. However,
using an anatomically realistic representation of rabbit ventricles, the
dynamics differed, matching what was recorded experimentally: with diacetyl monoxime, only a single
spiral wave corresponding to ventricular tachycardia was observed (Fig. 3A),
while with cytochalasin-D, an initiated spiral wave
broke apart into multiple waves (Fig. 3B), corresponding to ventricular
fibrillation. Analysis of these results showed that the cytochalasin-D
model produced more complex dynamics because, unlike the diacetyl-monoxime
model, spirals waves experienced tight turns, making them vulnerable to several
breakup mechanisms. Specifically, fibrillation arose from an
instability of the core induced by the anisotropic fiber rotation and by
dynamically created regions of dispersion of refractoriness produced by wave
front-back interaction due to the large size of the wavelength relative to the
ventricular size.

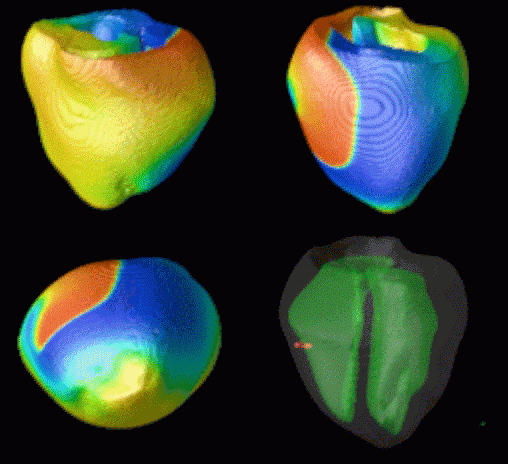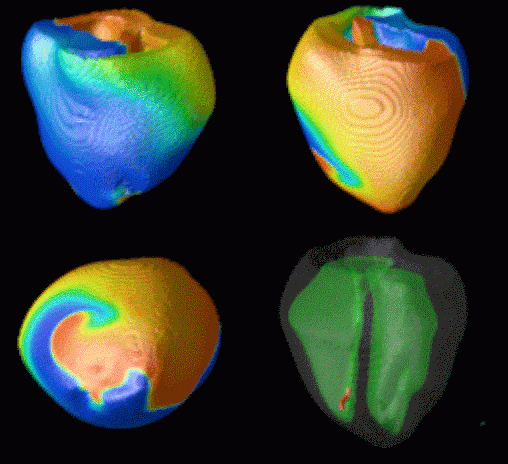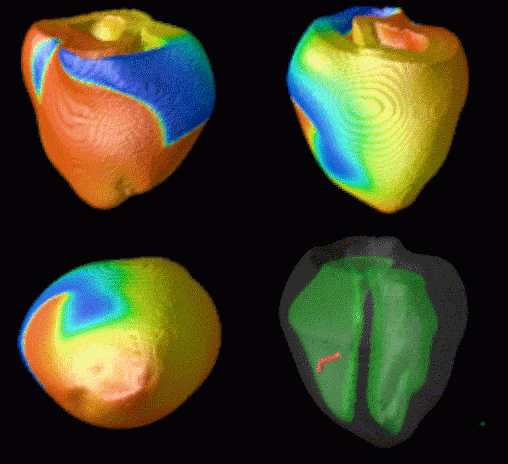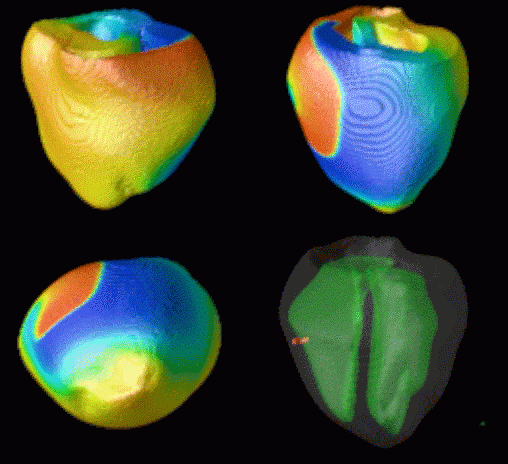
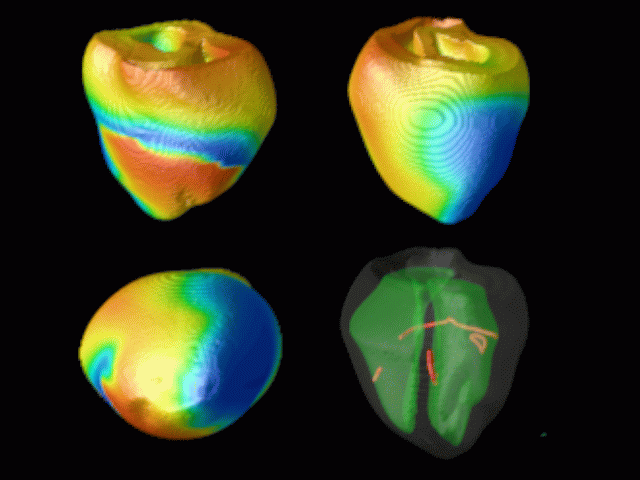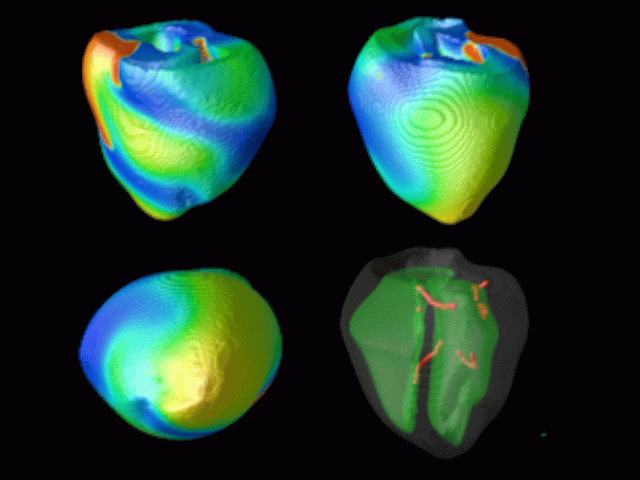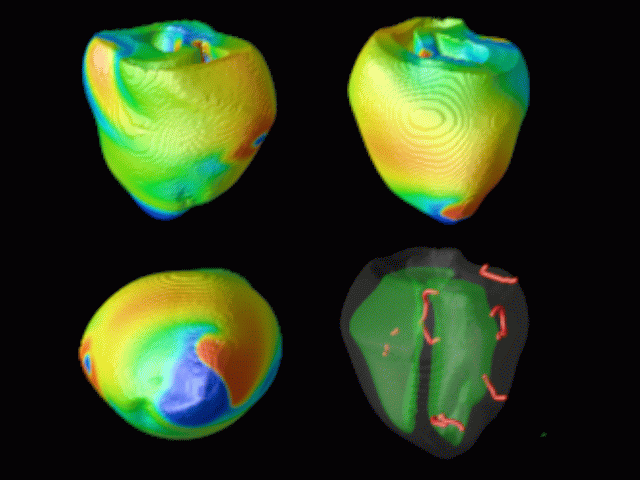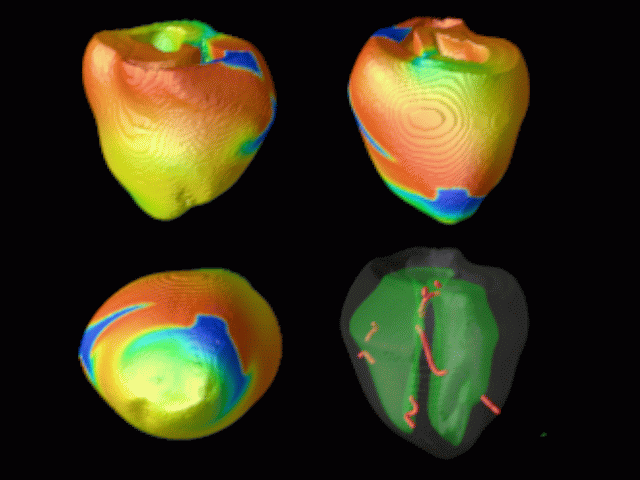
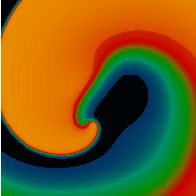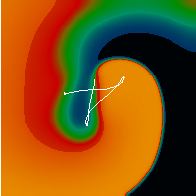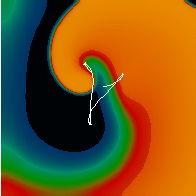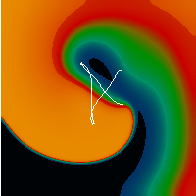
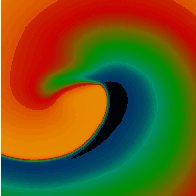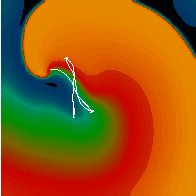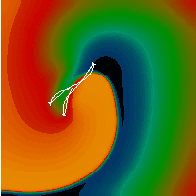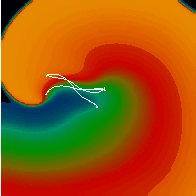
Figure 3. Simulated ventricular tachycardia
(A) and fibrillation (B) using an anatomically realistic model of the rabbit
ventricles including fiber orientation information. Electrical potential is
shown, with voltage ranging from –85 mV (blue) to 25 mV (red).
Anterior, posterior and apical views are shown, along with a semi-transparent
view showing the locations of transmural reentries
in red against the endocardial surfaces in green.
{kind=link}
{kind=link}
Movies of experimental ventricular tachycardia in the presence of diacetyl monoxime and ventricular fibrillation in the presence of cytochalasin-D.
{kind=link}
{kind=link}
Movies of stable spiral waves in two dimensions using models of rabbit ventricular electrophysiology in the presence of diacetyl monoxime and in the presence of cytochalasin-D.
{kind=link}
{kind=link}
Modeling normal canine
ventricular cell and tissue electrophysiology. Over the last decade, transmural gradients in
electrophysiological properties have been described. We have used
published experimental data in the canine heart to develop models of the three
dominant types of cells: epicardial, endocardial, and
specialized midmyocardial “M” cells. In addition, we subdivided the
canine ventricular anatomical model into spatial regions inhabited
by each of these cell types, as shown in Figure 4A for a representative cross-section.
Then we introduced a Purkinje network to reproduce a published activation
sequence observed experimentally (see Figure 4B) and simulated a series of
normal activations and the accompanying ECG. We compared the results with and
without the heterogeneity, and also with
and without an apex-base gradient in
repolarization. We found that although the apex-base gradient increased the
magnitude of an ECG deflection that indicates repolarization (the T-wave), the transmural heterogeneity was necessary for T-wave polarity
to be correct (upright), as shown in Figure 4C-D, in agreement with a current
hypothesis of the origin of the T-wave.

Figure 4. (A) Transmural heterogeneities in the canine ventricular
model. The red region shows M cells are assigned in the midmyocardium, with
endocardial cells in the remaining inside tissue and epicardial cells in the
remaining outside tissue. (B) Activation sequence in simulated canine
ventricles. The results agree well with the experimentally obtained
activation sequence of Durrer. (C) and (D)
Representative pseudo-ECGs obtained in simulations. Tissue is homogeneous in
(C) and includes transmural heterogeneity in (D),
whose upright deflection following each spike closely resembles healthy ECGs.
{kind=link}
Incorporating realistic intracellular calcium dynamics and contraction. Over the last several years, it has been demonstrated that voltage and calcium dynamics can operate independently, and recent optical mapping experiments have shown that the two systems sometimes desynchronize during fibrillation. Only one recently published mathematical model has been shown to allow independent behavior of the voltage and calcium systems at fast pacing rates. I have been working to couple this calcium model with a model of the cardiac action potential. Preliminary results are shown in Fig. 5, where alternans in voltage (A) are shown in concert with alternans in the intracellular calcium concentration (B).

Figure 5. Simulation of voltage
oscillations known as electrical alternans (A) driven by alternans in
intracellular calcium (B). Calcium alternans occur only at relatively fast rates
but generally disappear at extremely fast rates.
Back to Elizabeth Cherry's home page